Today is my last morning in Tulsa at SEAC 2017. I spent all day yesterday in the "Paleoindian and Early Archaic Southeast" symposium: 18 papers that included state-by-state updates of what we know and the data we have and treatments of topics such as megafauna in the Southeast, plant use by early foragers in the region, wet site archaeology in Florida, lithic technologies, etc. It was a marathon.
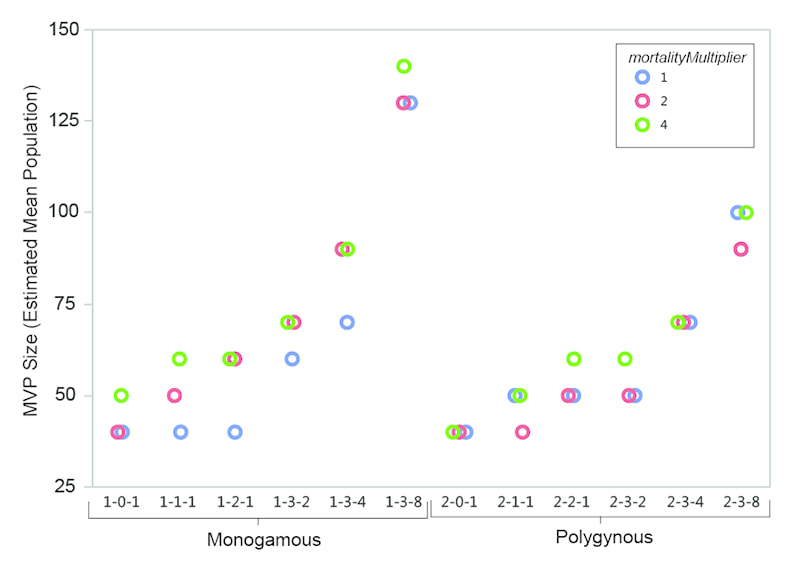
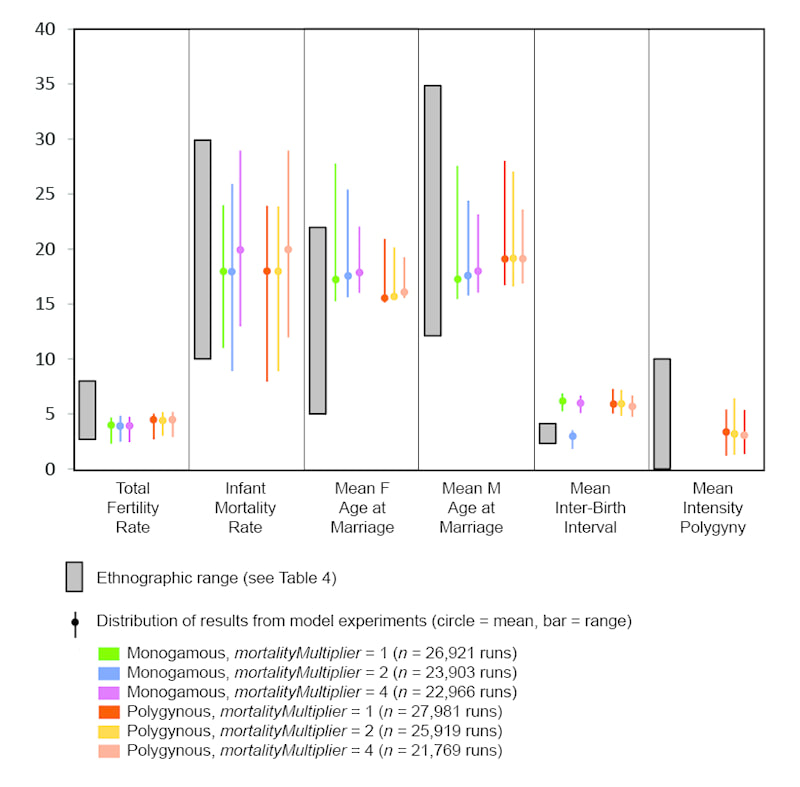
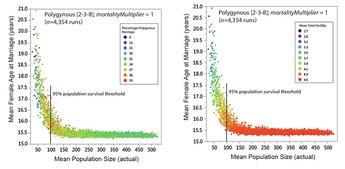
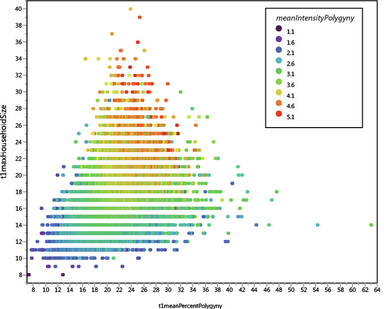
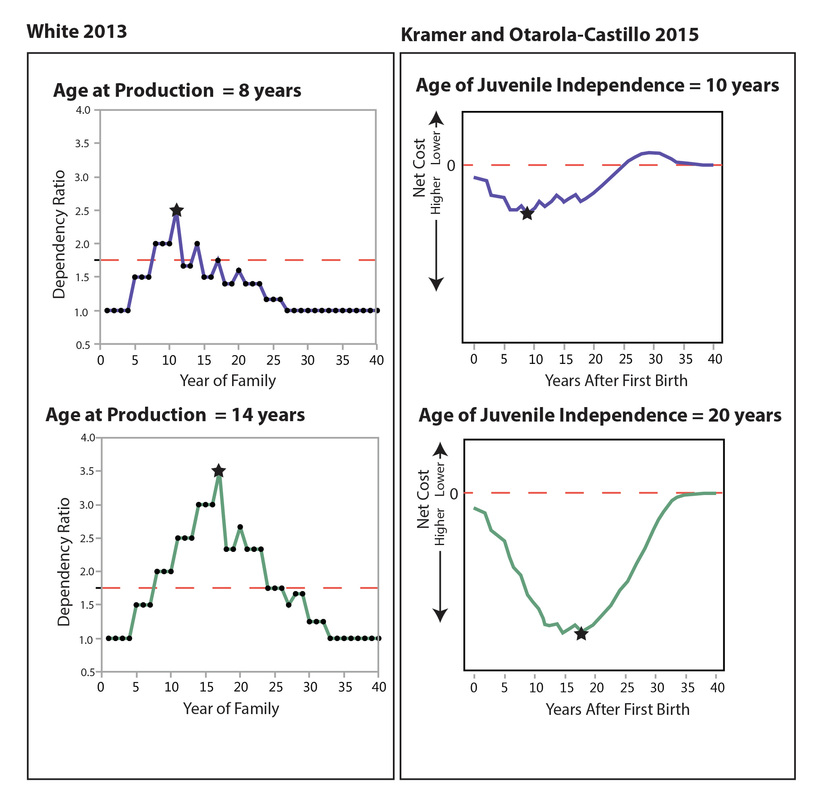
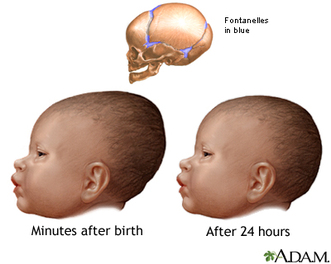
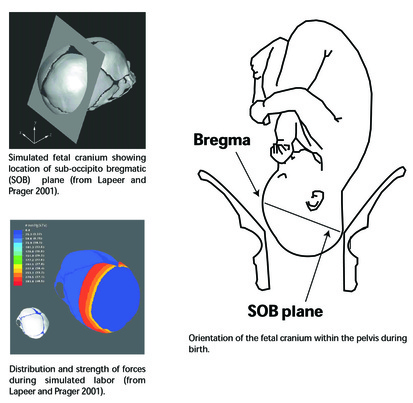
My presentation with David Anderson was last in the lineup. I was tasked with an effort at a "big picture" demography paper. It was a lot to talk about in a short time (20 minutes) -- a difficult balancing act to discuss the dense data from such a large area and be able to explain how I tried to integrate it all into a geographical/chronological model that can be evaluated on a region-by-region basis. Anyway . . . the detail will be there in the publications that result from the endeavor.
I uploaded a pdf of my presentation here. Some of the details will change as we work through the process of refining the analysis and dividing the content into multiple papers. But you should be able to get a decent idea of what we were going for.
My presentation with David Anderson was last in the lineup. I was tasked with an effort at a "big picture" demography paper. It was a lot to talk about in a short time (20 minutes) -- a difficult balancing act to discuss the dense data from such a large area and be able to explain how I tried to integrate it all into a geographical/chronological model that can be evaluated on a region-by-region basis. Anyway . . . the detail will be there in the publications that result from the endeavor.
I uploaded a pdf of my presentation here. Some of the details will change as we work through the process of refining the analysis and dividing the content into multiple papers. But you should be able to get a decent idea of what we were going for.



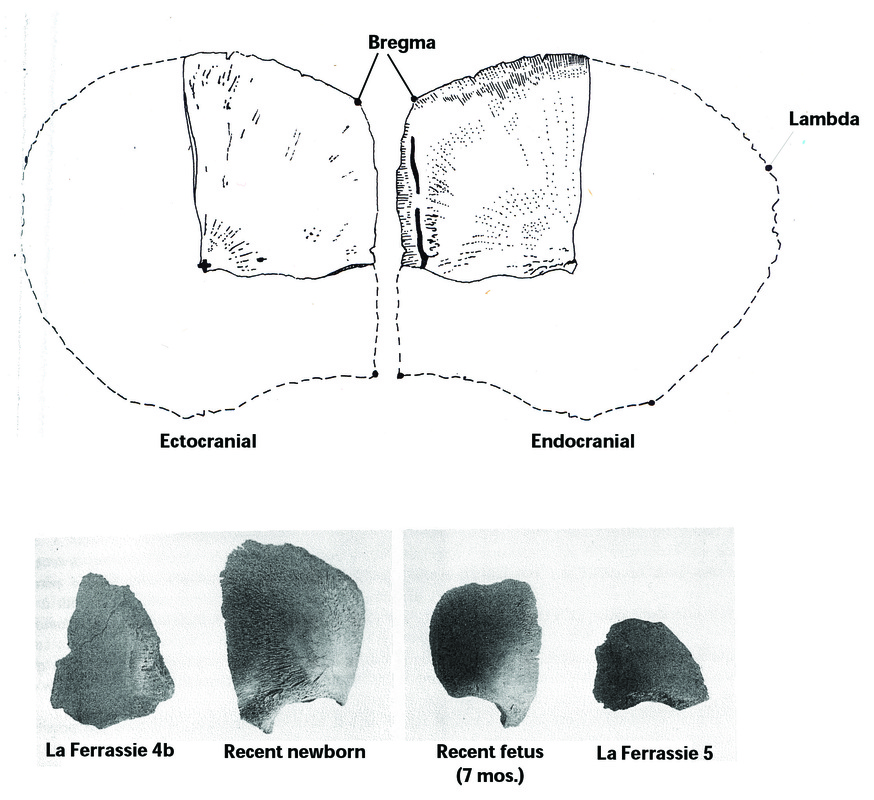
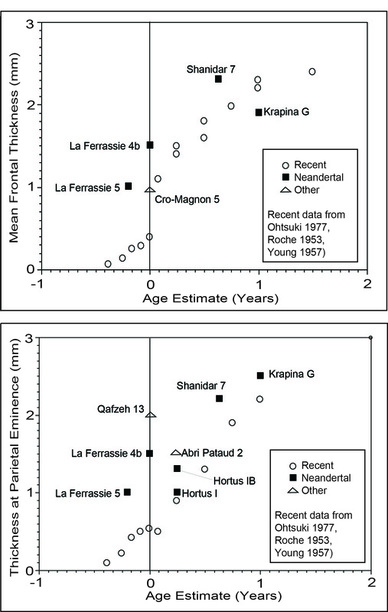
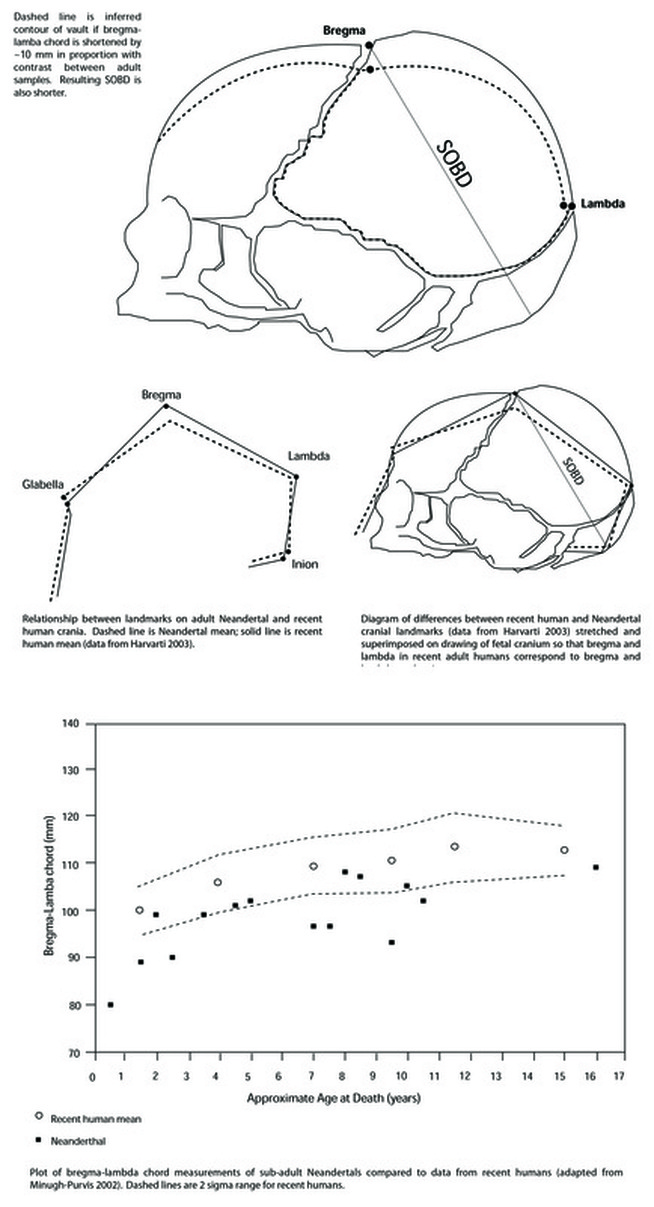


 RSS Feed
RSS Feed
