After having set computer modeling aside (out of necessity) for the past year, I'm ready to start getting back to that component of my research. I'm hoping to get things ramped back up this semester and be doing some fairly hardcore computational work by next semester. I've got some technical and logistical issues to address, and I'll surely have to spend some time getting back up to speed with Java and Repast. But it shouldn't be too bad
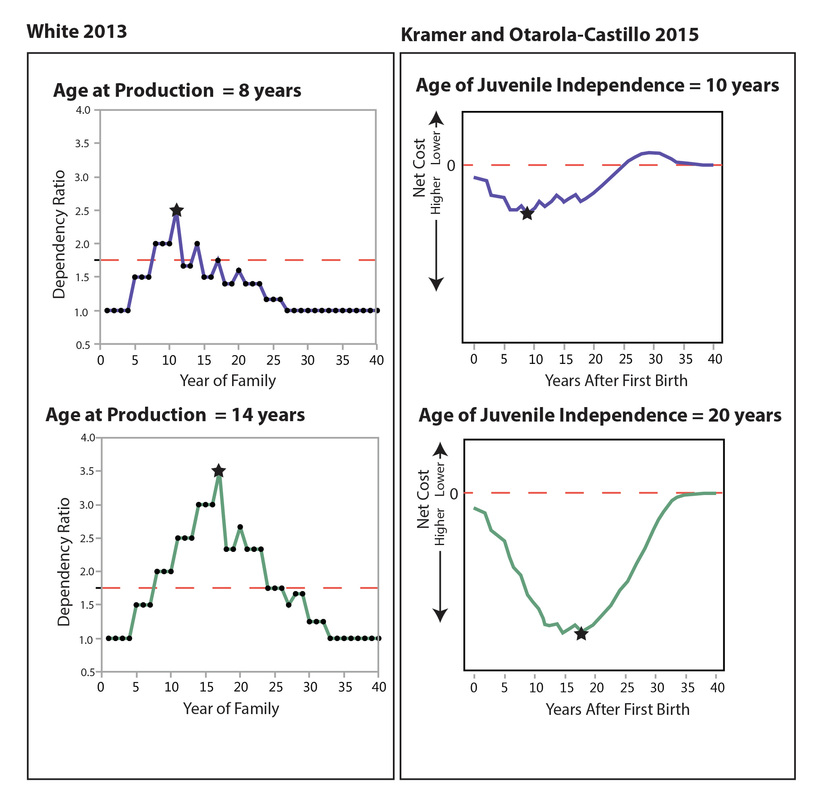
One of the things I'd like to do when I get up and running again is take another look at how the children's labor might affect the dependency ratio and household-level calculations about family size, etc. (see this paper for my first attempt to address the issue). In the models I'm currently using (e.g., the ForagerNet series), I represent a child as either a "producer" or a "non-producer" by comparing the child's age to the value of a parameter that specifies the age at which children become producers. Clearly that's a significant simplification of reality: children do not magically become "producers" overnight on their 8th birthdays or their 12th birthdays, etc.

I was watching my two boys (ages 4.7 years and 2.2 years) shell beans at the kitchen counter over the weekend, and it got me wondering what kind (if any) quantitative data are available about age-based changes in the proficiency of children in doing various kinds of productive tasks. Watching my two kids, there was a huge difference in the proficiency (and interest level) of the older and younger boys. The older one stuck with the task until we ran out of beans and, while not as fast as an adult, was really pretty good. Despite an equal contribution of energy (at least in the beginning), the productivity of the younger one was much lower. Part of that had to do with the desire to do a victory dance for each single bean that he managed to pull from a pod.
What would the age-based "proficiency curve" of bean processing look like? A linear progression? A rapid increase in proficiency between ages 2 and 6? At what age do individuals reach "full proficiency" in processing beans?
Are there ethnographic data available that would allow me to understand how proficiency at processing various kinds of plant foods changes with age? How about tasks related to gathering? Or planting?
There has been an increase in interest in the lives of children in hunting-gathering societies (see this paper by Nurit Bird-David), and I'm hoping that some quantitative data are available from recent ethnographic studies (e.g., see this 1994 paper by Blurton Jones et al., this 2004 paper by Raymond Hames and Patricia Draper, this 2009 volume edited by Barry Hewlett and Michael Lamb). I'm less familiar with possible sources for quantitative data relevant to modeling age-based changes in children's work proficiency in agricultural societies. Maybe there are government agencies or NGOs that monitor that sort of thing (the site of the International Labour Organization discusses children's domestic work, for example).
Anyway, this blog post is just a placeholder. I'm interested in tracking down some data if they're available (if you know of any, please let me know!).
If there are not suitable data already out there, I might have to generate some of my own through some controlled experimentation. Let me know if you like beans. We may soon be producing a surplus at my house. And I may soon ask to borrow the participation of children of various ages.
What would the age-based "proficiency curve" of bean processing look like? A linear progression? A rapid increase in proficiency between ages 2 and 6? At what age do individuals reach "full proficiency" in processing beans?
Are there ethnographic data available that would allow me to understand how proficiency at processing various kinds of plant foods changes with age? How about tasks related to gathering? Or planting?
There has been an increase in interest in the lives of children in hunting-gathering societies (see this paper by Nurit Bird-David), and I'm hoping that some quantitative data are available from recent ethnographic studies (e.g., see this 1994 paper by Blurton Jones et al., this 2004 paper by Raymond Hames and Patricia Draper, this 2009 volume edited by Barry Hewlett and Michael Lamb). I'm less familiar with possible sources for quantitative data relevant to modeling age-based changes in children's work proficiency in agricultural societies. Maybe there are government agencies or NGOs that monitor that sort of thing (the site of the International Labour Organization discusses children's domestic work, for example).
Anyway, this blog post is just a placeholder. I'm interested in tracking down some data if they're available (if you know of any, please let me know!).
If there are not suitable data already out there, I might have to generate some of my own through some controlled experimentation. Let me know if you like beans. We may soon be producing a surplus at my house. And I may soon ask to borrow the participation of children of various ages.




 RSS Feed
RSS Feed
