This morning's announcement of the first results from the Rising Star Expedition did not disappoint: a 35-page open access paper with 47 authors (Berger et al. 2015) describing fossil remains (Homo naledi) from at least 15 individuals recovered since 2013 from a cave in South Africa, and another paper by Dirks et al. (2015) describing the physical context of the fossils. I'm friends with several of the authors, and I am so happy for them both personally and professionally. And I'm jealous. But we'll leave that aside for now.
There are so many things that are awesome about this project that it's hard to even know where to start. If you don't follow paleoanthropology closely, you may not fully grasp how unusual this discovery is, how novel the approach to excavation and analysis was, and how @#!$&*% badass the results are. The story of the Rising Star Expedition is the Mad Max: Fury Road of paleoanthropology. I hope we start seeing more movies like this.
This is the largest single hominin fossil assemblage yet discovered in Africa, it is remarkable well-preserved, and it was analyzed and reported upon in record time. Two years from discovery to publication is blisteringly fast in the world of paleoanthropology (the publication of Ardipethecus ramidus, a 4.4 million-year-old putative hominin from Ethiopia, famously took over 15 years). Speeding up initial analysis by inviting a crowd of young, hungry scholars to contribute to the work (hence the 47 authors on the first paper) was a masterful stroke by project leader Lee Berger. The Rising Star Expedition has convincingly demonstrated the utility of a new paradigm that challenges traditional notions about how paleoanthropology should be done and what constitutes an appropriate pace for analysis and publication. Bravo on that front: totally @#!$&*% badass.
There are so many things that are awesome about this project that it's hard to even know where to start. If you don't follow paleoanthropology closely, you may not fully grasp how unusual this discovery is, how novel the approach to excavation and analysis was, and how @#!$&*% badass the results are. The story of the Rising Star Expedition is the Mad Max: Fury Road of paleoanthropology. I hope we start seeing more movies like this.
This is the largest single hominin fossil assemblage yet discovered in Africa, it is remarkable well-preserved, and it was analyzed and reported upon in record time. Two years from discovery to publication is blisteringly fast in the world of paleoanthropology (the publication of Ardipethecus ramidus, a 4.4 million-year-old putative hominin from Ethiopia, famously took over 15 years). Speeding up initial analysis by inviting a crowd of young, hungry scholars to contribute to the work (hence the 47 authors on the first paper) was a masterful stroke by project leader Lee Berger. The Rising Star Expedition has convincingly demonstrated the utility of a new paradigm that challenges traditional notions about how paleoanthropology should be done and what constitutes an appropriate pace for analysis and publication. Bravo on that front: totally @#!$&*% badass.
The initial findings/interpretations/conclusions from Rising Star are also pretty @#!$&*% badass. Here are a few that jump out to me as important during a first quick pass through the paper:
Mosaic of Primitive and Derived Features. According to the authors, the skeletons of Homo naledi preserve a mixture of human-like and australopithecine-like features unlike that seen in any other fossil remains:
H. naledi has humanlike manipulatory adaptations of the hand and wrist. It also exhibits a humanlike foot and lower limb. These humanlike aspects are contrasted in the postcrania with a more primititive or australopith-like trunk, shoulder, pelvis and proximal femur (Berger et al. 2015:2).
Mosaic of Primitive and Derived Features. According to the authors, the skeletons of Homo naledi preserve a mixture of human-like and australopithecine-like features unlike that seen in any other fossil remains:
H. naledi has humanlike manipulatory adaptations of the hand and wrist. It also exhibits a humanlike foot and lower limb. These humanlike aspects are contrasted in the postcrania with a more primititive or australopith-like trunk, shoulder, pelvis and proximal femur (Berger et al. 2015:2).
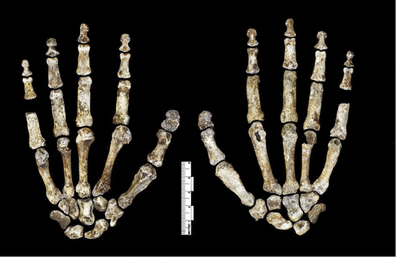 Two views of the hand of Homo naledi (Figure 6 from Berger et al. 2015). The hand was discovered still articulated.
Two views of the hand of Homo naledi (Figure 6 from Berger et al. 2015). The hand was discovered still articulated.
The appearance of a different "mosaic" of primitive and derived features was also described by Lee Berger for the Australopithecus sediba fossils (a rather late gracile australopithecine from South Africa), highlighting the difficulty understanding extinct hominins based on single bones or very fragmentary skeletons. The skeleton of sediba appeared to have such a strange mixture of features that some researchers suggested the remains were actually those of two different species mixed together. I think it will be pretty difficult to make that argument for the naledi remains because of the number of specimens and their apparent morphological homogeneity. The implications of the mosaic aspects of the Rising Star fossils will have to be dealt with rather than dismissed. The features that we associate with being human apparently did not come together as a "package deal."
South, Not East? Identifying naledi as a member of our genus means it is a potential human ancestor. The center of gravity in the study of the emergence of the genus Homo has been in East Africa for many decades. But if naledi is ancestral to our lineage, what does that mean for the fossils of possible human ancestors found in East Africa? Do they get pushed out of the lineage? Do they have to leave the adult table at Thanksgiving and go sit with the little kids? There is no possible fossil assemblage from East Africa of early Homo that compares with the one from Rising Star in terms of size and its potential to tell us about so many aspects of the skeleton. I think that means that, unless you want to just assume that East Africa is where everything important happened, naledi has to be dealt with in any serious discussion of the origins of Homo. And that brings us once again to the "species" issue.
What Does "Species" Mean? I wrote a little bit about species concepts inthis post from May when a new species (Australopithecus deyiremeda) was proposed for the fossil of another purported human ancestor. My point in that post was that if we're imagining that each "species" is a reproductively isolated population, naming a new species has a lot of implications that I think are tough to justify based on fragmentary fossils. It's most problematic when species are named based on very little physical evidence. There's a lot of material from Rising Star, however, and I would guess that many people are comfortable with creating a new taxon based on the amount of material that Berger et al. (2015) have analyzed.
That's fine. But I'm still wondering what is meant by the term "species." I didn't see a definition of "species" in the paper (it's possible I missed it), but I did see this passage in the National Geographic story about Rising Star that came out today:
"Berger himself thinks the right metaphor for human evolution, instead of a tree branching from a single root, is a braided stream: a river that divides into channels, only to merge again downstream. Similarly, the various hominin types that inhabited the landscapes of Africa must at some point have diverged from a common ancestor. But then farther down the river of time they may have coalesced again, so that we, at the river’s mouth, carry in us today a bit of East Africa, a bit of South Africa, and a whole lot of history we have no notion of whatsoever."
I like the metaphor of a braided stream, but it sounds a lot more like we're describing populations within a single biological species rather than species-level divergences. Once the "species" streams diverge through reproductive isolation, how could they ever converge again? Using a biological species concept, they can't. We may need to give things names so we can talk about them and compare them, but it seems to me, again, that our taxonomic terminology and practices are ill-suited and maybe counter-productive for actually describing and understanding the patterns and processes that we're interested in.
But There's No Date! There are no dates associated with the Rising Star fossils. That means we really have no independent handle on where in time they go, which means it's hard to use them to test specific hypotheses about the patterns, processes, and history of human evolution. Without dates it will really tough to understand how they might fit in with the East African materials, or sediba, or the recently-announced 3.3 million-year-old stone tools from Kenya. Wherever these fossils "go" in time they'll make a splash, but we don't know right now where the ripples will be. I hope we get some information about that soon.
Was naledi Burying Its Dead? Surely one of the most controversial interpretations of the Rising Star assemblage will be that it accumulated through intentional disposal of the dead. That's the conclusion of this paper by Dirks et al. (2015):
"Preliminary evidence is consistent with deliberate body disposal in a single location, by a hominin species other than Homo sapiens, at an as-yet unknown date."
That lack of a date is especially painful here. The earliest claims that I know of for regularized treatment of the dead come from the site of Sima de los Huesos (Atapuerca), where the bodies of 28 individuals were thrown into a cave in Spain around 400-500 thousand years ago. Evidence for the patterned, ritualized treatment of the dead at Atapuerca fits well with other possible signs of a cognitive emergence about half a million years ago, including the shell of similar age announced last year that was apparently carved by Homo erectus and evidence of the removal of flesh from the Bodo cranium at about 600,000 years ago.
If the Rising Star fossils date to the origins of Homo, as the researchers suggest, and if the assemblage accumulated through intentional cultural behavior, it will push ritual treatment of the dead much farther back in time than we have ever considered. Two million years ago? I bet there are probably people out there who still don't even think Neanderthals were burying their dead. That's sure to spark some arguments.
Jealousy aside, it's going to be great to watch what happens next. I'm sure there will be more results from Rising Star (hopefully soon), and I'm sure there will be a lot of reaction to what came out today. It's a totally @#!$&*% badass project with totally @#!$&*% badass results. This is a great time to be paying attention to human evolution - maybe the best ever. Fingers crossed we get an announcement of australopithecine DNA soon.
South, Not East? Identifying naledi as a member of our genus means it is a potential human ancestor. The center of gravity in the study of the emergence of the genus Homo has been in East Africa for many decades. But if naledi is ancestral to our lineage, what does that mean for the fossils of possible human ancestors found in East Africa? Do they get pushed out of the lineage? Do they have to leave the adult table at Thanksgiving and go sit with the little kids? There is no possible fossil assemblage from East Africa of early Homo that compares with the one from Rising Star in terms of size and its potential to tell us about so many aspects of the skeleton. I think that means that, unless you want to just assume that East Africa is where everything important happened, naledi has to be dealt with in any serious discussion of the origins of Homo. And that brings us once again to the "species" issue.
What Does "Species" Mean? I wrote a little bit about species concepts inthis post from May when a new species (Australopithecus deyiremeda) was proposed for the fossil of another purported human ancestor. My point in that post was that if we're imagining that each "species" is a reproductively isolated population, naming a new species has a lot of implications that I think are tough to justify based on fragmentary fossils. It's most problematic when species are named based on very little physical evidence. There's a lot of material from Rising Star, however, and I would guess that many people are comfortable with creating a new taxon based on the amount of material that Berger et al. (2015) have analyzed.
That's fine. But I'm still wondering what is meant by the term "species." I didn't see a definition of "species" in the paper (it's possible I missed it), but I did see this passage in the National Geographic story about Rising Star that came out today:
"Berger himself thinks the right metaphor for human evolution, instead of a tree branching from a single root, is a braided stream: a river that divides into channels, only to merge again downstream. Similarly, the various hominin types that inhabited the landscapes of Africa must at some point have diverged from a common ancestor. But then farther down the river of time they may have coalesced again, so that we, at the river’s mouth, carry in us today a bit of East Africa, a bit of South Africa, and a whole lot of history we have no notion of whatsoever."
I like the metaphor of a braided stream, but it sounds a lot more like we're describing populations within a single biological species rather than species-level divergences. Once the "species" streams diverge through reproductive isolation, how could they ever converge again? Using a biological species concept, they can't. We may need to give things names so we can talk about them and compare them, but it seems to me, again, that our taxonomic terminology and practices are ill-suited and maybe counter-productive for actually describing and understanding the patterns and processes that we're interested in.
But There's No Date! There are no dates associated with the Rising Star fossils. That means we really have no independent handle on where in time they go, which means it's hard to use them to test specific hypotheses about the patterns, processes, and history of human evolution. Without dates it will really tough to understand how they might fit in with the East African materials, or sediba, or the recently-announced 3.3 million-year-old stone tools from Kenya. Wherever these fossils "go" in time they'll make a splash, but we don't know right now where the ripples will be. I hope we get some information about that soon.
Was naledi Burying Its Dead? Surely one of the most controversial interpretations of the Rising Star assemblage will be that it accumulated through intentional disposal of the dead. That's the conclusion of this paper by Dirks et al. (2015):
"Preliminary evidence is consistent with deliberate body disposal in a single location, by a hominin species other than Homo sapiens, at an as-yet unknown date."
That lack of a date is especially painful here. The earliest claims that I know of for regularized treatment of the dead come from the site of Sima de los Huesos (Atapuerca), where the bodies of 28 individuals were thrown into a cave in Spain around 400-500 thousand years ago. Evidence for the patterned, ritualized treatment of the dead at Atapuerca fits well with other possible signs of a cognitive emergence about half a million years ago, including the shell of similar age announced last year that was apparently carved by Homo erectus and evidence of the removal of flesh from the Bodo cranium at about 600,000 years ago.
If the Rising Star fossils date to the origins of Homo, as the researchers suggest, and if the assemblage accumulated through intentional cultural behavior, it will push ritual treatment of the dead much farther back in time than we have ever considered. Two million years ago? I bet there are probably people out there who still don't even think Neanderthals were burying their dead. That's sure to spark some arguments.
Jealousy aside, it's going to be great to watch what happens next. I'm sure there will be more results from Rising Star (hopefully soon), and I'm sure there will be a lot of reaction to what came out today. It's a totally @#!$&*% badass project with totally @#!$&*% badass results. This is a great time to be paying attention to human evolution - maybe the best ever. Fingers crossed we get an announcement of australopithecine DNA soon.
Berger, L., Hawks, J., de Ruiter, D., Churchill, S., Schmid, P., Delezene, L., Kivell, T., Garvin, H., Williams, S., DeSilva, J., Skinner, M., Musiba, C., Cameron, N., Holliday, T., Harcourt-Smith, W., Ackermann, R., Bastir, M., Bogin, B., Bolter, D., Brophy, J., Cofran, Z., Congdon, K., Deane, A., Dembo, M., Drapeau, M., Elliott, M., Feuerriegel, E., Garcia-Martinez, D., Green, D., Gurtov, A., Irish, J., Kruger, A., Laird, M., Marchi, D., Meyer, M., Nalla, S., Negash, E., Orr, C., Radovcic, D., Schroeder, L., Scott, J., Throckmorton, Z., Tocheri, M., VanSickle, C., Walker, C., Wei, P., & Zipfel, B. (2015). , a new species of the genus from the Dinaledi Chamber, South Africa eLife, 4 DOI: 10.7554/eLife.09560








 RSS Feed
RSS Feed
